Post provided by Dr Andrés Baselga

A key property of biodiversity is that it is not evenly distributed around the world. In other words, different sites are usually home to different biological communities. Quantifying the differences among biological communities is a major step towards understanding how and why biodiversity is distributed in the way it is.
The term beta diversity was introduced by R.H. Whittaker in 1960. He defined it as “the extent of change in community composition, or degree of community differentiation, in relation to a complex-gradient of environment, or a pattern of environments”. In his original paper, Whittaker proposed several ways to quantify beta diversity. In its simplest form (which we will call strict sense or multiplicative beta diversity), beta diversity is defined as the ratio between gamma (regional) and alpha (local) diversities (Whittaker, 1960; Jost, 2007). Therefore, it is the effective number of distinct compositional units in the region (Tuomisto, 2010). Essentially, beta diversity quantifies the number of different communities in the region. So it’s clear that beta diversity does not only account for the relationship between local and regional diversity, but also informs about the degree of differentiation among biological communities. This is because alpha and gamma diversities are different if (and only if) the biological communities within the region are different.
It’s easy to demonstrate how beta diversity varies from the minimum to the maximum differentiation of local assemblages in a region. For simplicity, we will quantify biological diversity as species richness (number of species), but it’s important to remember that alpha, beta and gamma diversities can also be defined to account for richness and relative abundances (see Jost, 2007 for a detailed explanation). When local assemblages are all identical (minimum differentiation), alpha diversity equals gamma diversity, and beta diversity equals 1 (figure below).

When the mean local species richness (alpha diversity) equals the regional species richness (gamma diversity), their ratio (beta diversity) equals unity. This means that in our first example region (above) there is only one distinct compositional unit (i.e. only one “community”).
When local assemblages are all completely different (maximum differentiation), gamma diversity equals the multiplication of alpha diversity by the number of sites (N). So beta diversity equals N, meaning that in our second example region there are N distinct compositional units, so N different “communities” (figure below).

Standardizing Beta Diversity
As seen above, multiplicative beta diversity (gamma/alpha) ranges from 1 to N (number of sites in the region). Therefore, to get a measure of differentiation independent of the number of sites (N) involved in the calculation, we need to standardize beta diversity. For example, the Sørensen index of dissimilarity is just beta-1 divided by N-1, which ranges between 0 and 1, and is independent of the number of sites (N). In general, dissimilarity indices that are monotonic transformations of strict sense beta diversity (for example the Sørensen and Jaccard indices) are appropriate measures of differences among biological communities (see Chao et al., 2012).
What does “Different” Mean?
The meaning of “difference” applied to biological communities is not unidimensional. The first thought we all have when thinking about beta diversity, or difference between biological assemblages, is the replacement of some species by others.
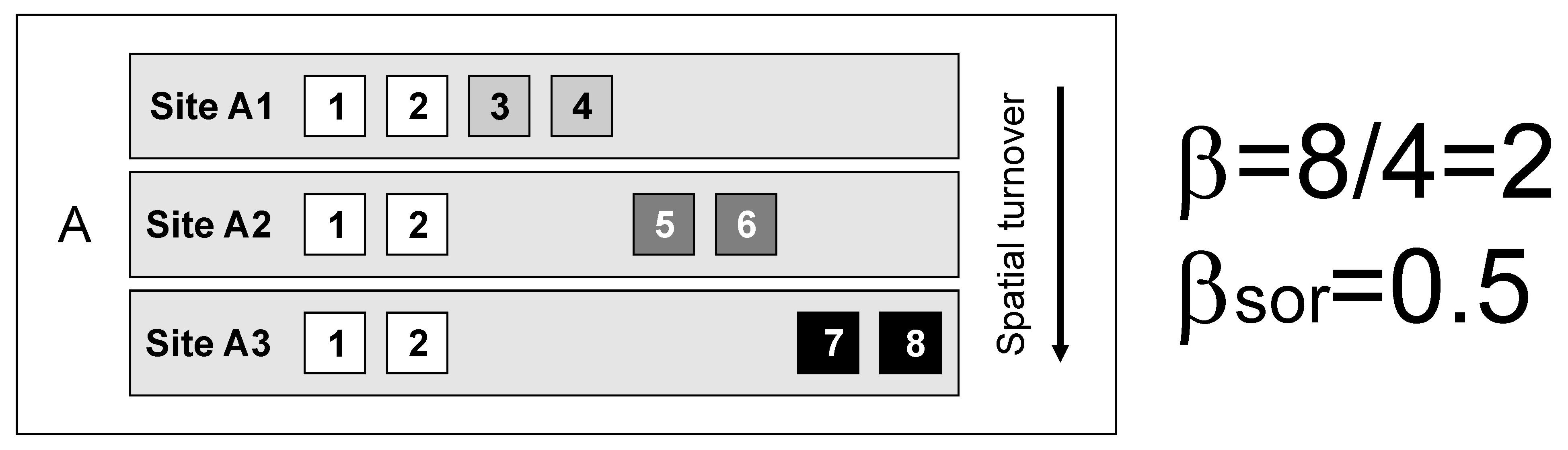
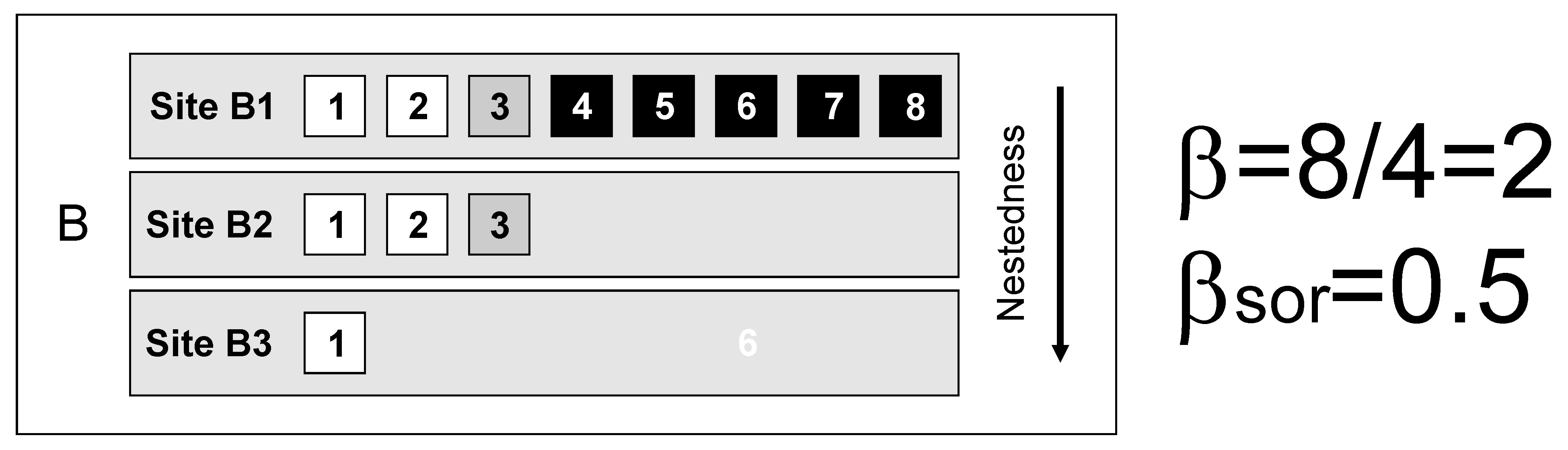
However, the nested loss of species from the richest to the poorest locality can also cause alpha and gamma diversity to differ. This has the potential to yield the same value of beta diversity for strikingly different patterns.
Of course, the observed patterns can be a combination of both replacement and species loss.
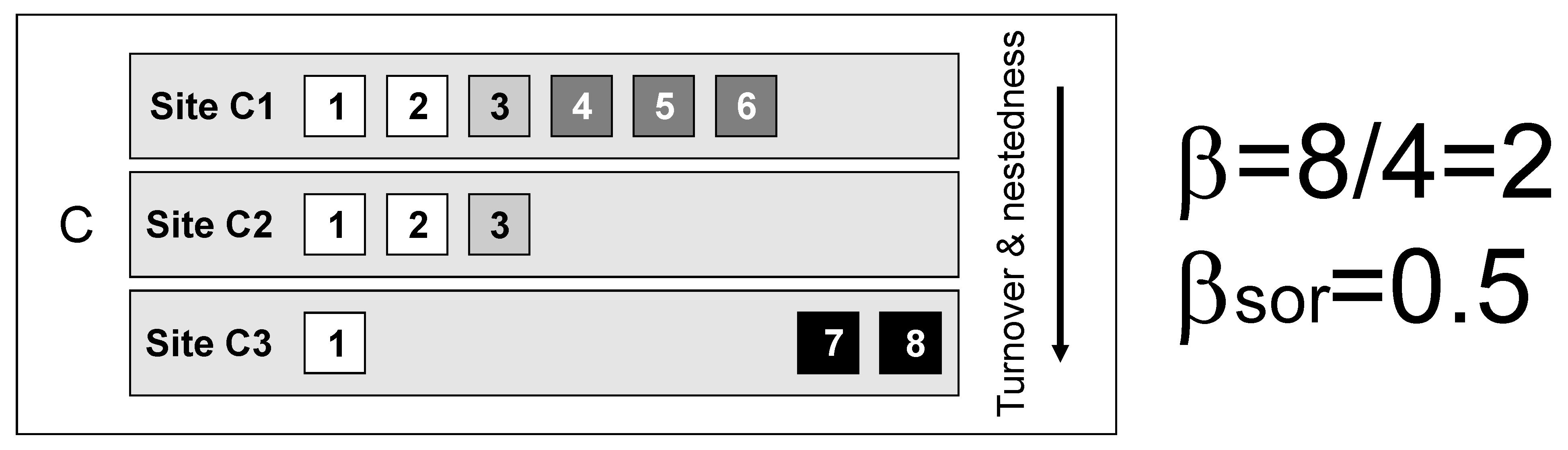
In the three situations above (A-C), gamma diversity (8 species) and alpha diversity (mean site diversity = 4 species) are identical, so multiplicative beta diversity (gamma/alpha) and the related dissimilarity indices (e.g., Sørensen, Jaccard) also have identical values.
How can Species Replacement and Loss be Separated?
The Simpson index of dissimilarity (Simpson, 1943; Simpson, 1960) was intended to remove the effects of richness difference in communities. In fact, it considers nested assemblages to be perfectly similar and provides the replacement component of beta diversity. In the absence of richness difference, Simpson and Sørensen indices have identical values, meaning that their difference accounts for the nestedness-resultant component of dissimilarity. This leads to a framework in which total dissimilarity (Sørensen or Jaccard indices) can be additively portioned into replacement and nestedness-resultant components (see Baselga, 2010 and Baselga, 2012 for a detailed explanation). The same approach can be used to separate components of abundance-based dissimilarity (Baselga, 2013; Legendre, 2014), functional dissimilarity (Villeger et al., 2013) and phylogenetic dissimilarity (Leprieur et al., 2012). Functions to compute all of these dissimilarity indices are available in R package betapart (see Baselga & Orme, 2012).
Besides these extensions of the partitioning method, an alternative partitioning framework has also been proposed (Podani & Schmera, 2011 and extensions). This is intended to separate the replacement and richness-difference components of dissimilarity. Pierre Legendre has recently reviewed the interpretation of both portioning frameworks, clarifying, among many other things, the conceptual differences between the nestedness-resultant and richness-different components of alternative methods (Legendre, 2014). Even more recently, Fabien Leprieur and I have systematically compared the behaviour of parallel components in both alternative partitioning methods (Baselga & Leprieur, 2015). We concluded that the replacement components of both alternative methods account for different concepts and that only the replacement component of the original framework (Baselga, 2010; Baselga, 2012) is independent of richness difference. However, the parallel component in the alternative framework (Podani & Schmera, 2011; Carvalho et al., 2012) is mathematically constrained by richness difference. As a result, only the original framework (Baselga, 2010; Baselga, 2012, and extensions) allows for the separation of (i) the patterns of assemblage differentiation derived from species replacement which is not mathematically constrained by species richness differences from (ii) the patterns of assemblage differentiation in nested subsets.
References
Baselga, A. (2010) Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography, 19, 134-143.
Baselga, A. (2012) The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Global Ecology and Biogeography, 21, 1223-1232.
Baselga, A. (2013) Separating the two components of abundance-based dissimilarity: balanced changes in abundance vs. abundance gradients. Methods in Ecology and Evolution, 4, 552–557
Baselga, A. & Leprieur, F. (2015) Comparing methods to separate components of beta diversity. Methods in Ecology and Evolution, in press.
Baselga, A. & Orme, C. D. L. (2012) betapart: an R package for the study of beta diversity. Methods in Ecology and Evolution, 3, 808-812.
Carvalho, J. C., Cardoso, P. & Gomes, P. (2012) Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Global Ecology and Biogeography, 21, 760–771.
Chao, A., Chiu, C.-H. & Hsieh, T. C. (2012) Proposing a resolution to debates on diversity partitioning. Ecology, 39, 2037-2051.
Jost, L. (2007) Partitioning diversity into independent alpha and beta components. Ecology, 88, 2427-2439.
Legendre, P. (2014) Interpreting the replacement and richness difference components of beta diversity. Global Ecology and Biogeography, 23, 1324–1334.
Leprieur, F., Albouy, C., De Bortoli, J., Cowman, P. F., Belwood, D. R. & Mouillot, D. (2012) Quantifying phylogenetic beta diversity: distinguishing between ‘true’ turnover of lineages and phylogenetic diversity gradients. PLoS One, 7, e42760.
Podani, J. & Schmera, D. (2011) A new conceptual and methodological framework for exploring and explaining pattern in presence – absence data. Oikos, 120, 1625-1638.
Simpson, G. G. (1943) Mammals and the Nature of Continents. American Journal of Science, 241, 1-31.
Simpson, G. G. (1960) Notes on the measurement of faunal resemblance. American Journal of Science, 258, 300-311.
Tuomisto, H. (2010) A diversity of beta diversities: straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography, 33, 2-22.
Villeger, S., Grenouillet, G. & Brosse, S. (2013) Decomposing functional β-diversity reveals that low functional β-diversity is driven by low functional turnover in European fish assemblages. Global Ecology and Biogeography, 22, 671–681.
Whittaker, R. H. (1960) Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs, 30, 280-338.

A really worth to share material! Really useful and well assembled! Thanks
Excellent! Helped a lot, thanks.
This is, by far, the best basic explanation of beta/alpha/gamma diversity I’ve ever read.
Really useful and easy to understand concepts.
Incredibly helpful! Thanks so much!