Post provided by ROB LEWIS & MEELIS PÄRTEL
Our understanding of how biological diversity works has been advanced by a long history of observing species and linking patterns to ecological processes. However, we generally don’t focus as much on those species that aren’t observed, or in other words ‘absent species’. But, can absent species provide valuable information?
Dark diversity – a set of species absent from a particular site but which belong to its species pool – has the potential to be as ecologically meaningful as observed diversity. Part of the species pool concept, understanding dark diversity is relatively straightforward.
The Basic Theory of Dark Diversity
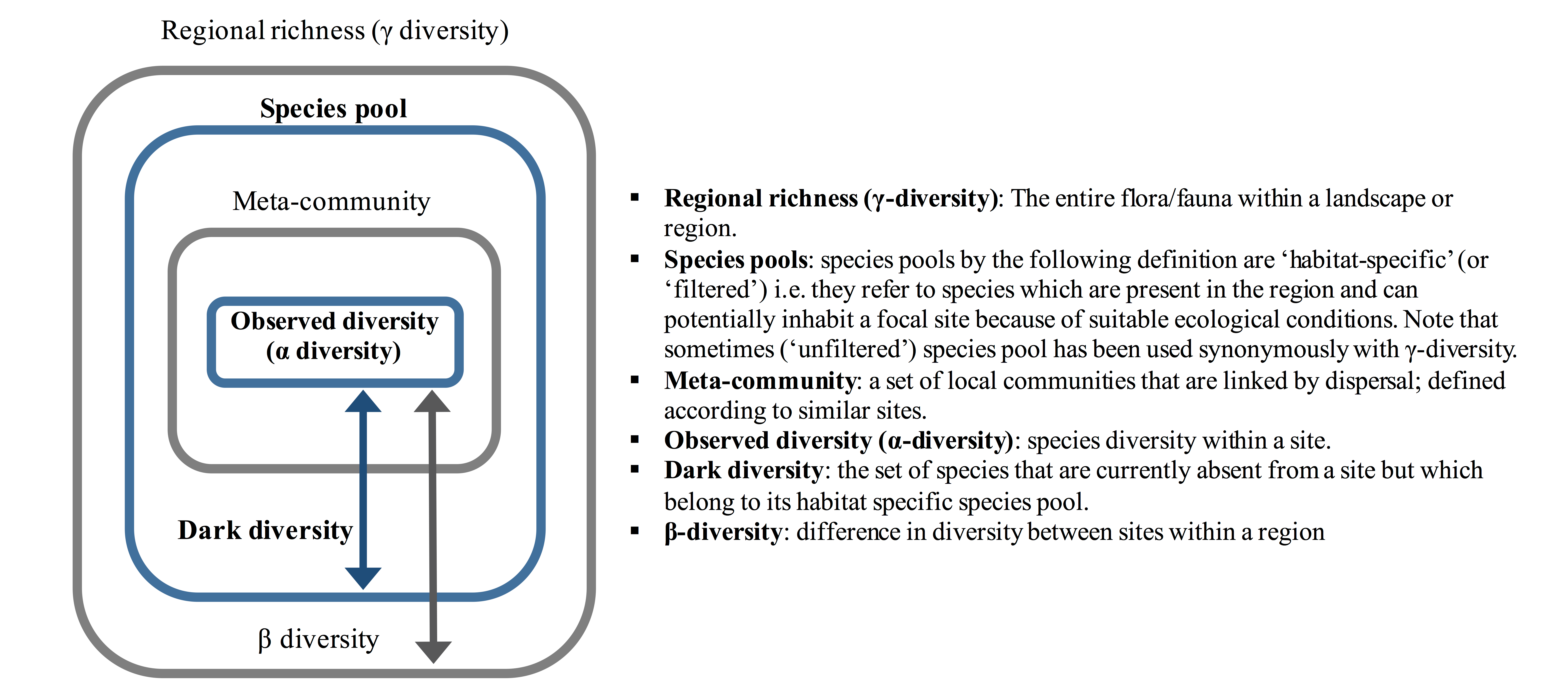
To begin learning about dark diversity, there are two important terms that we need to define: ‘species pool’ and ‘focal community’. A ‘species pool’ is a set of species present in a particular region or landscape that can potentially inhabit a particular observed community because of suitable local ecological conditions.
A ‘focal community’ is the set of species that have been observed in a particular region or landscape (this is the ‘observed community’ and can also be referred to as alpha diversity). For a given focal community to become established, the species within it must have overcome dispersal pressures as well as environmental and biotic filters.

Species present in the surrounding region or landscape are no exception – they must have overcome these factors to establish successful populations. The species observed in any location are therefore always a subset of the total number of species belonging to a particular species pool.
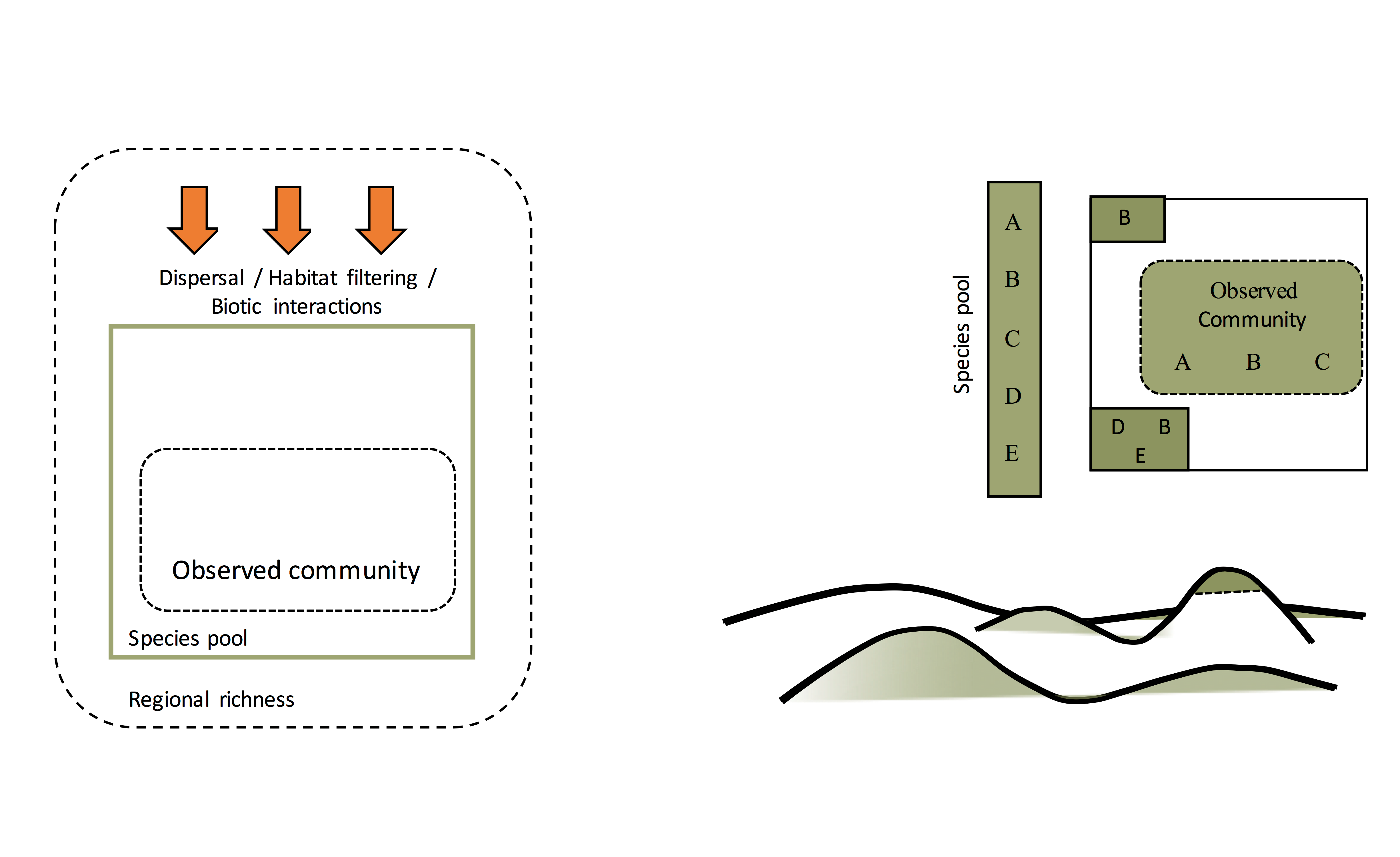
Dark diversity is the portion of the species pool not observed in the focal community. Simply, it constitutes species that are present in the surrounding landscape or region that can potentially colonize the site in question, but are currently absent from it.
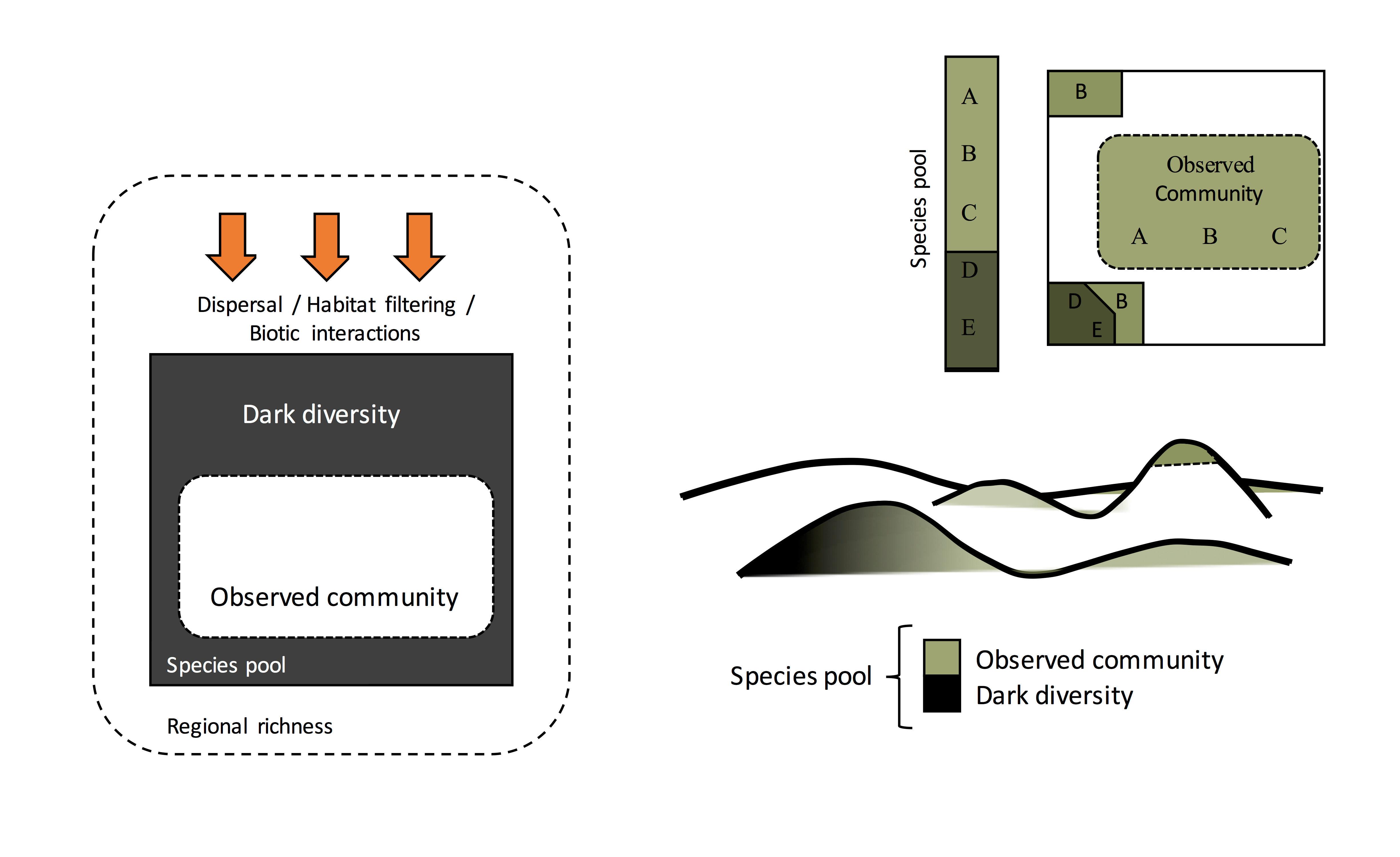
The figures above (A-C), show a hypothetical focal community in an alpine grassland. Here the species pool is made up of species A-E, the observed community species A-C, and by deduction its dark diversity species D & E.
It’s useful to note that dark diversity can be applied at any spatial scale (e.g. 1m x 1m or 50km x 50km), and can be used for the study of plants, animals, insects, fungi or microbes. It can also be used to derive a relativised index of biodiversity termed ‘completeness’, (i.e. how complete a habitat/region is relative to its respective species pool), which allows for biodiversity comparisons among dissimilar communities within or across regions and/or habitat types.
Dark Diversity NOT Beta Diversity
Species pools by definition are habitat-specific (‘filtered’)– they are ‘a suite of species which are always related to a certain community type’. However, the term ‘species pool’ has been used synonymously with gamma diversity (the entire list of species across a landscape or region that is likely to encompass multiple habitat types, without regard for any abiotic habitat filter). The use of the term ‘species pool’ in this way cannot be used to study dark diversity.
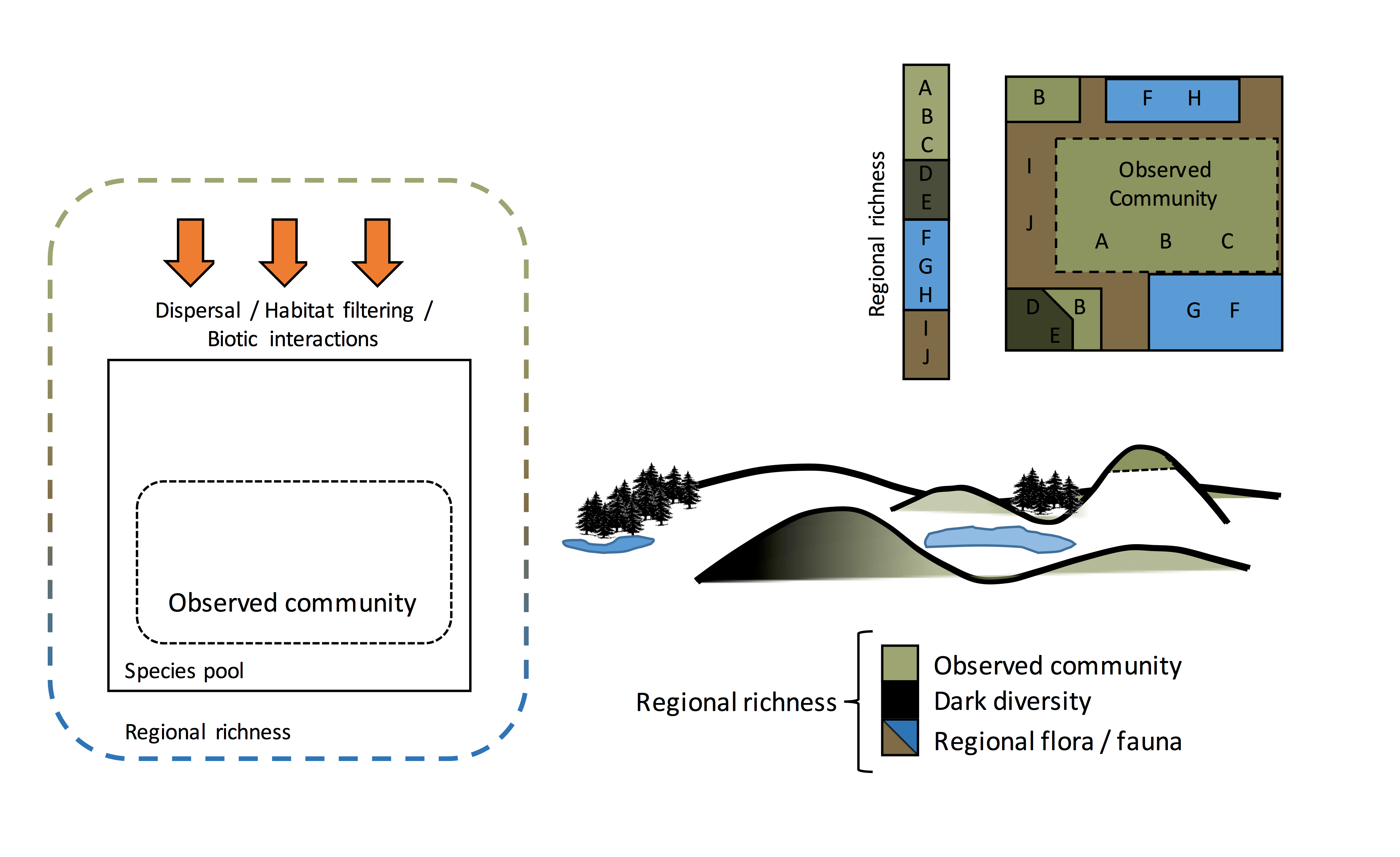
Across environmental gradients (e.g. moisture, temperature, exposure), multiple biological communities exist (e.g. forests, wetlands and grasslands; Figure D above). The species within them constitute the entire regional flora or fauna (regional richness), expressing a wide range of ecological requirements. For this reason, misuse of the term ‘species pool’ to constitute regional richness does not allow for estimating dark diversity. Depending on the habitat heterogeneity of the study area, any compositional differences between the observed community (strictly alpha diversity) and regional richness (strictly gamma diversity) would constitute the study of beta diversity and not dark diversity. For example, a species of trout may be a part of the regional richness of the example region above, but it would not be part of the species pool for the alpine grassland areas because environmental factors (lack of water) mean that it cannot survive there.
Measuring the Un-Measurable
The term ‘dark diversity’ deliberately draws on parallels with dark matter. Like dark matter, dark diversity cannot be seen nor directly measured. However, dark diversity can be accurately estimated and using readily available data.
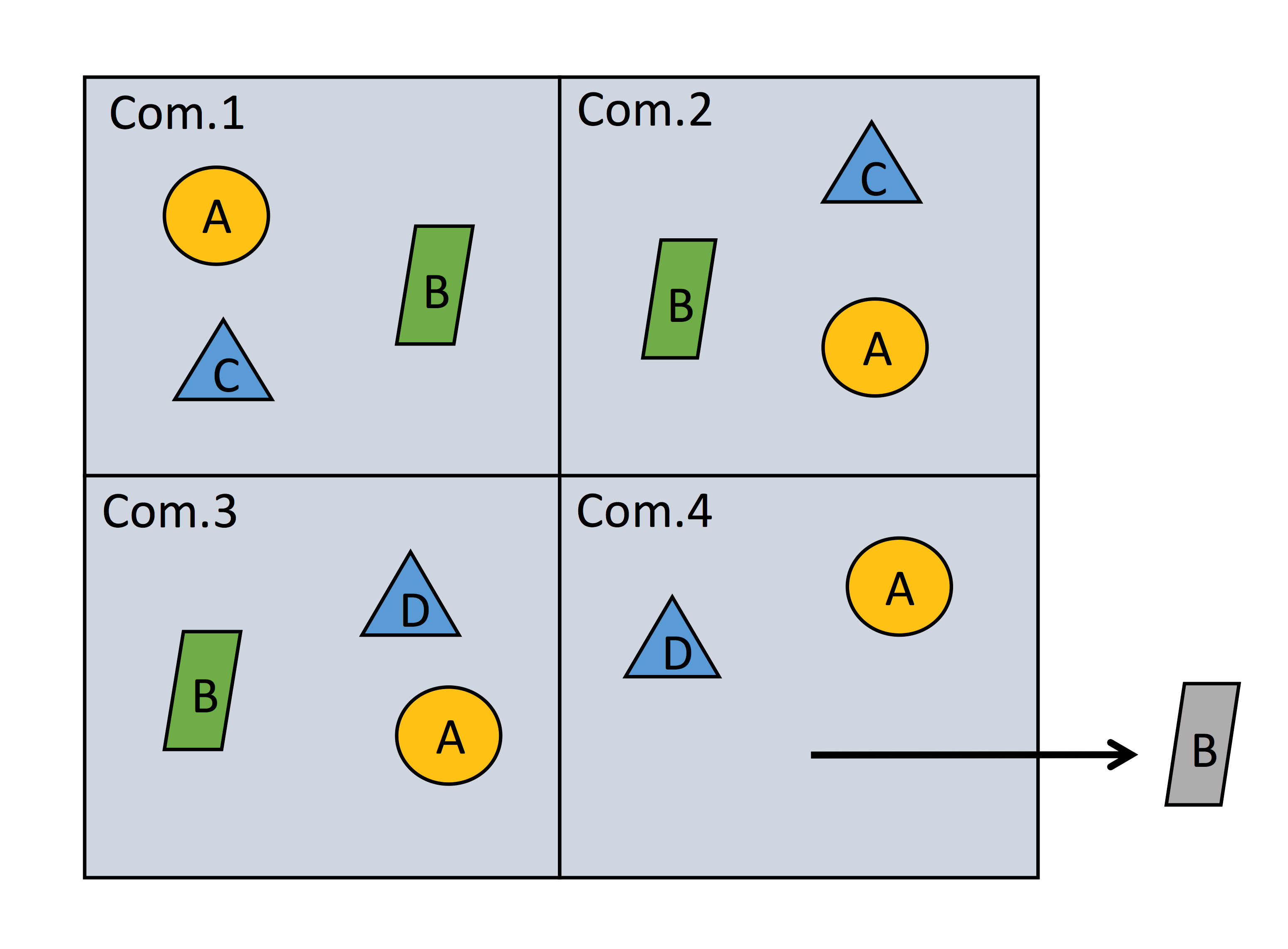
In an article recently published in Methods in Ecology and Evolution, we demonstrate how species co-occurrence patterns can be used for estimating dark diversity. Quantifying patterns of association between species over space (which helps to gauge the likelihood of a given species co-existing with another) are used to assign an absent species a probability value of co-existing among observed species for a given community. Probabilities of co-occurrence can then be used to compose a community’s dark diversity with reasonable accuracy. For details on how co-occurrence patterns can be used to compose dark diversity see Fig. 2 of Estimating dark diversity and species pools: an empirical assessment of two methods.
Application of Dark Diversity
Observable patterns provide only a portion of the information necessary for fully understanding the mechanisms and processes acting upon a particular population or community. Quantifying and understanding patterns of dark diversity provides additional and valuable information. A recent publication in Conservation Biology explores how the dark diversity concept can be used to address fundamental challenges faced by conservationists. One example focuses on restoration ecology, suggesting that an understanding of a restoration site’s dark diversity is helpful for several reasons. These include:
- Allowing for the comparison of a ‘restoration’ site species pool to a ‘reference’ site species pool.
- If the two species pools are very different, restoration success is unlikely to be effective in the short-term, or at least without intervention (e.g. species translocation; biomass removal).
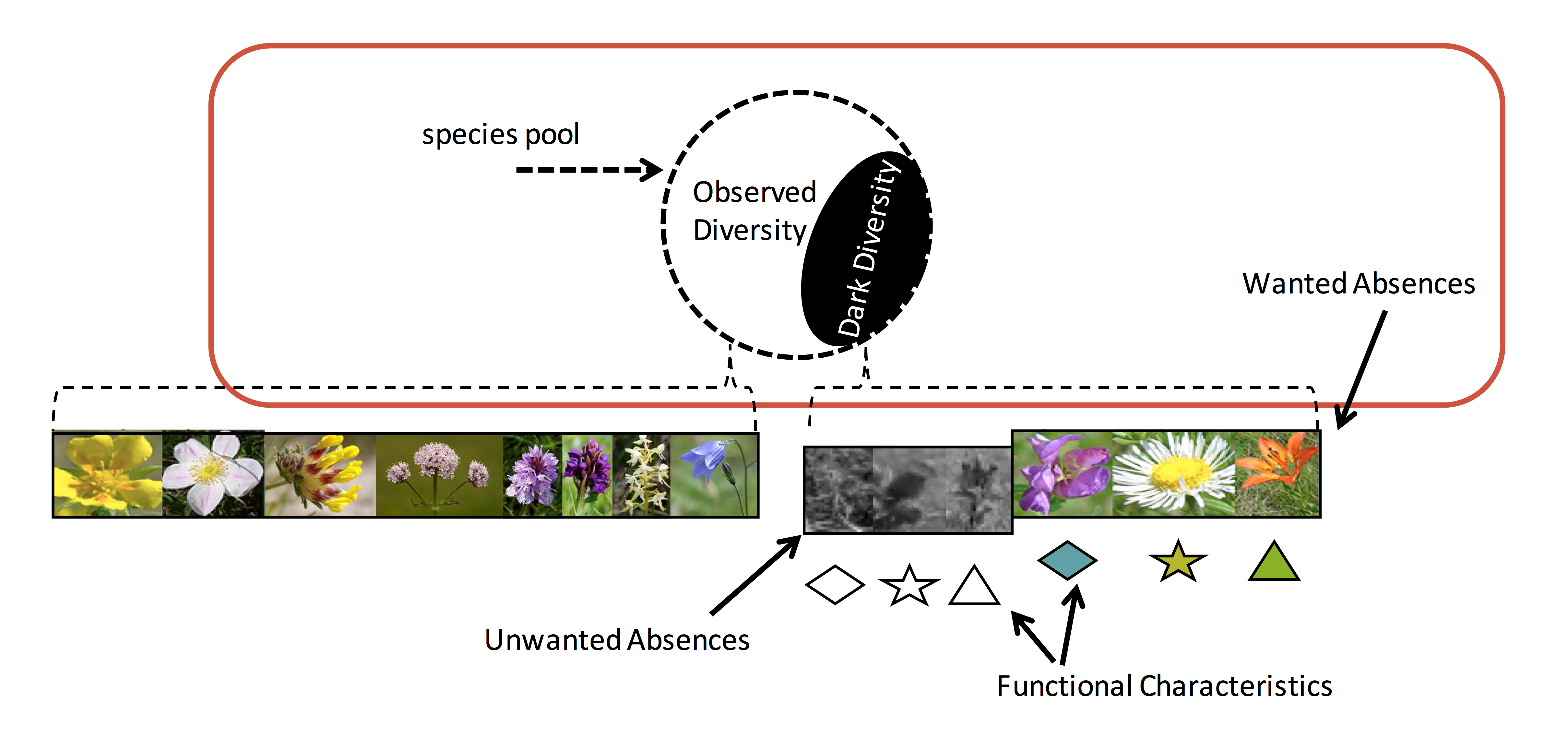
- Helping researchers to explore the functional characteristics of the absent species .
- The functional traits (i.e. a suite of characteristics that define how a species interacts with its environment and other species) of absent species can reveal clues as to why species may be absent (e.g. competition, predation, dispersal limitation), helping devise successful restoration management plans.
- The identification of ‘unwanted’ species.
- Not all species are desired (invasives, for example). Knowledge of potential ‘unwanted’ species belonging to a restoration site’s dark diversity can help develop and implement preventative measures to ensure these species remain part of the dark diversity.
There are many reasons to expect an understanding of dark diversity to contribute towards understanding patterns of biological diversity. Implemented more widely, the study of dark diversity can complement existing ecological approaches and concepts (e.g. meta-community dynamics, beta diversity), bringing together additional ecological information for describing study systems. Current advancements in methods are promising and we hope to see further development in the future.
To find out more about dark diversity and to learn how to estimate it, read the Methods in Ecology and Evolution article ‘Estimating dark diversity and species pools: an empirical assessment of two methods’.
