Blog escrito por: gustavo burin
This post is also available in English

Há alguns dias, me deparei com um interessante vídeo sobre os chamados “fósseis vivos”. O vídeo focou mais nos problemas de usá-los como argumentos contra a teoria da evolução, e aproveitei a oportunidade para falar mais sobre essas linhagens longevas.
‘Fóssil vivo‘ é um termo usado para descrever linhagens que acredita-se terem se originado há muito tempo e que mantêm características que se assemelham a seus parentes fósseis. Alguns exemplos bem conhecidos dessas linhagens são os Tuatara da Nova Zelândia (Sphenodon punctatus) e as árvores Gingkos (Gingko biloba).
Como podemos identificar um ‘fóssil vivo’?
Geralmente essas linhagens apresentam ramos muito longos nas filogenias moleculares, muito mais longos do que as linhagens atuais normalmente mostram (veja exemplo abaixo). O comprimento destes ramos é proporcional ao tempo (geralmente na escala dos milhões de anos). Estes ramos longos podem nos dar a impressão de que essas linhagens estão vivas desde centenas de milhões de anos atrás.

Mas, na realidade, elas são provavelmente muito, muito mais jovens do que isso. Esses ramos muito longos são provavelmente artefatos da maneira como essas filogenias são construídas. Para entender melhor isso, precisamos parar e revisar como uma filogenia molecular é construída.
Filogenias moleculares, na maioria das vezes, são construídas usando apenas dados de DNA de espécies existentes (já que é quase impossível obter informações confiáveis de DNA de fósseis). Como em qualquer outro estudo em ecologia e evolução, para reconstruir uma filogenia confiável precisamos de informação para o maior número de espécies possível. Do contrário, tanto as relações evolutivas entre as espécies quanto o tempo de divergência pode ser inferido incorretamente e todas as conclusões baseadas na filogenia não seriam confiáveis. Se o número de espécies com dados moleculares disponíveis for amplo o suficiente (correspondendo a uma fração significativa da riqueza total do grupo), os modelos de evolução molecular tentarão dizer como as sequências de DNA de todas essas espécies evoluíram a partir do ancestral comum mais recente (ACMR) de todo o grupo.
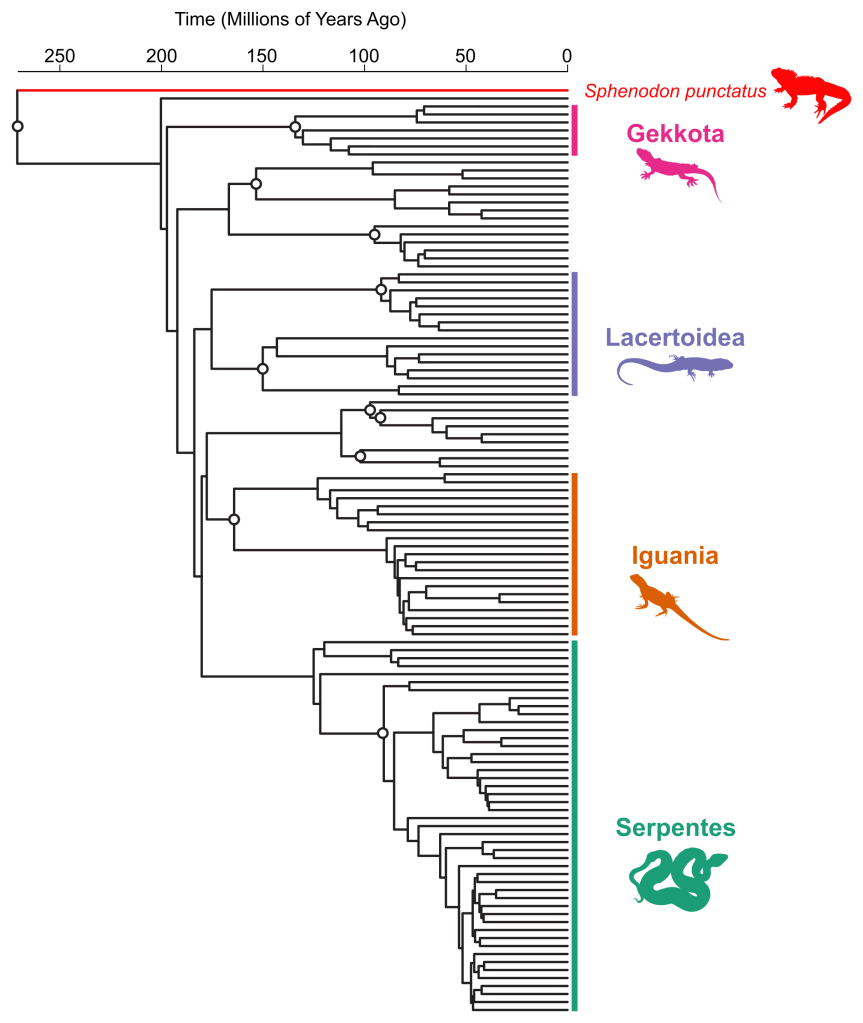
Os fósseis vivos pertencem à linhagens que divergiram há muito tempo da linhagem de ‘fósseis não vivos’, mas que possuem muito poucas espécies existentes. Portanto, esperamos que o DNA deles reflita essa história evolutiva e que as linhagens dos fósseis vivos sejam ligados à base da filogenia. Consequentemente, essas linhagens são representadas como tendo se originado há muito tempo, com ramos muito longos (até vários milhões de anos no caso da Tuatara, por exemplo). No entanto, é importante ter em mente uma limitação importante das filogenias moleculares: elas não registram informações diretas sobre a extinção de linhagens!
Como mencionei antes, filogenias moleculares são construídas usando informações de espécies existentes apenas (embora informações oriundas de fósseis são usadas para calibração do tempo). Isso significa que as linhagens originadas e extintas não serão representadas na filogenia resultante. Essa ausência dá a impressão de que nada acontecia o tempo todo que o fóssil vivo ‘existia’.
São fósseis vivos ‘espécies Highlander’?

Sabemos pelo registro fóssil que a duração média das espécies de mamíferos, por exemplo, é de cerca de 2 a 4 milhões de anos (embora isso varie bastante entre grupos diferentes, mas vamos usar esse valor como um referencial). Com isso em mente, somos praticamente forçados a perguntar “o que raios os fósseis vivos têm de diferente que lhes permitem viver 20 vezes mais do que uma espécie típica? Seriam eles o elo perdido que liga os vertebrados típicos à linhagem dos MacLeod?” A resposta para essas perguntas pode estar na extinção.
Explico: sabemos que longos ramos nas filogenias moleculares podem ser gerados através de longas histórias de extinção. Estes eventos apagam as informações sobre espécies que se originaram e se extinguiram ao longo da história evolutiva do grupo.
Vamos usar algumas simulações de processos macroevolutivos para ilustrar isso: ambas as filogenias mostradas na figura abaixo são representações da mesma simulação. A filogenia à esquerda contém todas as espécies (incluindo as extintas). A filogenia da direita, no entanto, contém apenas as espécies existentes. Podemos notar que alguns dos ramos parecem muito mais longos na filogenia à direita.

Ao olhar para esses ramos longos, nossa intuição nos faz pensar (pelo menos em um primeiro momento) que nada aconteceu ao longo deles. No entanto, com base em nossa compreensão da dinâmica evolutiva, o mais correto é esperar que de fato muitas coisas possam ter acontecido, com muitas espécies diferentes sendo geradas nesse meio tempo. A maioria dessas espécies, no entanto, foi extinta e apenas uma (ou poucas) permanece viva hoje para ‘contar a história’ de toda a linhagem. Devido à essa história de especiação e extinção, é provável que as espécies que ainda estão vivas hoje não tenham se originado muitos milhões de anos atrás, mas muito mais recentemente.
Esse é mais um motivo pelo qual o termo ‘fóssil vivo’ é muito pouco preciso. Além da dinâmica de diversificação, outra explicação possível para a semelhança entre as espécies existentes e seus parentes fósseis pode estar nas baixas taxas de evolução morfológica. Mas isso é assunto para outra hora.
Se você quiser conversar mais sobre macroevolução, me siga no twitter!
Para saber mais, leia os Virtual Issues da Methods in Ecology and Evolution ‘Advances in Phylogenetic Methods’ e ‘Phylogenetics and Comparative Methods’.

One thought on “O Problema com os ‘Fósseis Vivos’: Uma Perspectiva Filogenética Molecular”