Robert May Prize Shortlisted Article
Post provided by Claudio S. Quilodrán and Ashley T. Sendell-Price

Each year Methods in Ecology and Evolution awards the Robert May Prize to the best paper in the journal by an author at the start of their career. Claudio S. Quilodrán has been shortlisted for his article ‘The multiple population genetic and demographic routes to islands of genomic divergence’. In this blog, Claudio and co-author Ashley T. Sendell-Price discuss how their paper came to be and how their individual‐based simulation can be used to explore the dynamics of diverging genomes.
Imagine a songbird hatching on a small island in the Pacific Ocean. There is little chance that this bird will survive to sexual maturity and reproduce. However, if it does, this bird will contribute to the genetic diversity of the following generation, and potentially have an influence on the population’s evolutionary trajectory.
The way that populations diverge over time, potentially resulting in the emergence of new species, is a major topic in biology. This has motivated many generations of scientists, including Charles Darwin and Alfred Wallace, when they jointly proposed the theory of natural selection more than 160 years ago. Molecular tools developed during the last half-century have given us an improved understanding of the genetic basis of the speciation process. Genomic information from an increasing number of species has accumulated rapidly over the past few decades. This now includes genomic data from non-model species, such as birds hatching on small islands of the Pacific.
These empirical observations have shown that the genomes of divergent populations do not change uniformly, as some genomic regions differentiate at higher rates than others. Much effort has been applied to understanding why patterns of differentiation vary across the genome. However, reconstructing the evolutionary processes and understanding the extent to which these processes can generate recognizable signals at the level of the genome has remained challenging.
Evolutionary trajectories
One of the main challenges of analysing genomic or phenotypic patterns from empirical observations is that they result from a single evolutionary trajectory. Yet, similar patterns can be obtained from different processes that have influenced distinct evolutionary trajectories.

For instance, we can still imagine that our bird reaches sexual maturity and reproduces, bringing genetic novelties that influence a longer beak on its descendants. A motivated scientist that goes sampling on this island, some generations later, may observe most individuals with a longer beak. When comparing them with populations from surrounding islands, the scientist may argue that individuals with a longer beak are somehow better adapted to this island. A paper reviewer may also consider they survived by chance. Both options are likely, but since we have a single evolutionary trajectory observed from the field, deciding between them is an extremely difficult task. However, a main interest in simulating evolution is the opportunity of recreating evolutionary trajectories multiple times. This may help to depict the impact of contingency events, such as a storm that destroyed all of the nests on our island, with the exception of the one in which our bird was nesting. Multiple simulations have the potential to narrow the range of options that explain past evolutionary trajectories. Our bird may reborn at each simulation run.
The ‘glads’ R package
We present a flexible R package to simulate the evolution of diverging genomes (glads: genomic landscape of divergence simulations). The R package considers the life history of individuals with their genetic or genomic information to explore plausible evolutionary scenarios that could produce observed genomic divergence patterns. We use this tool to explore various evolutionary processes that may result in various levels of differentiation across the genome of diverging populations.
This package provides a novel method for projecting eco-evolutionary dynamics by considering the genomic information of individuals, with ecological interactions at the level of populations, communities and potentially ecosystems. This modelling framework may help to understand past evolutionary histories, as well as potential routes of future evolution.
The genomic islands of divergence
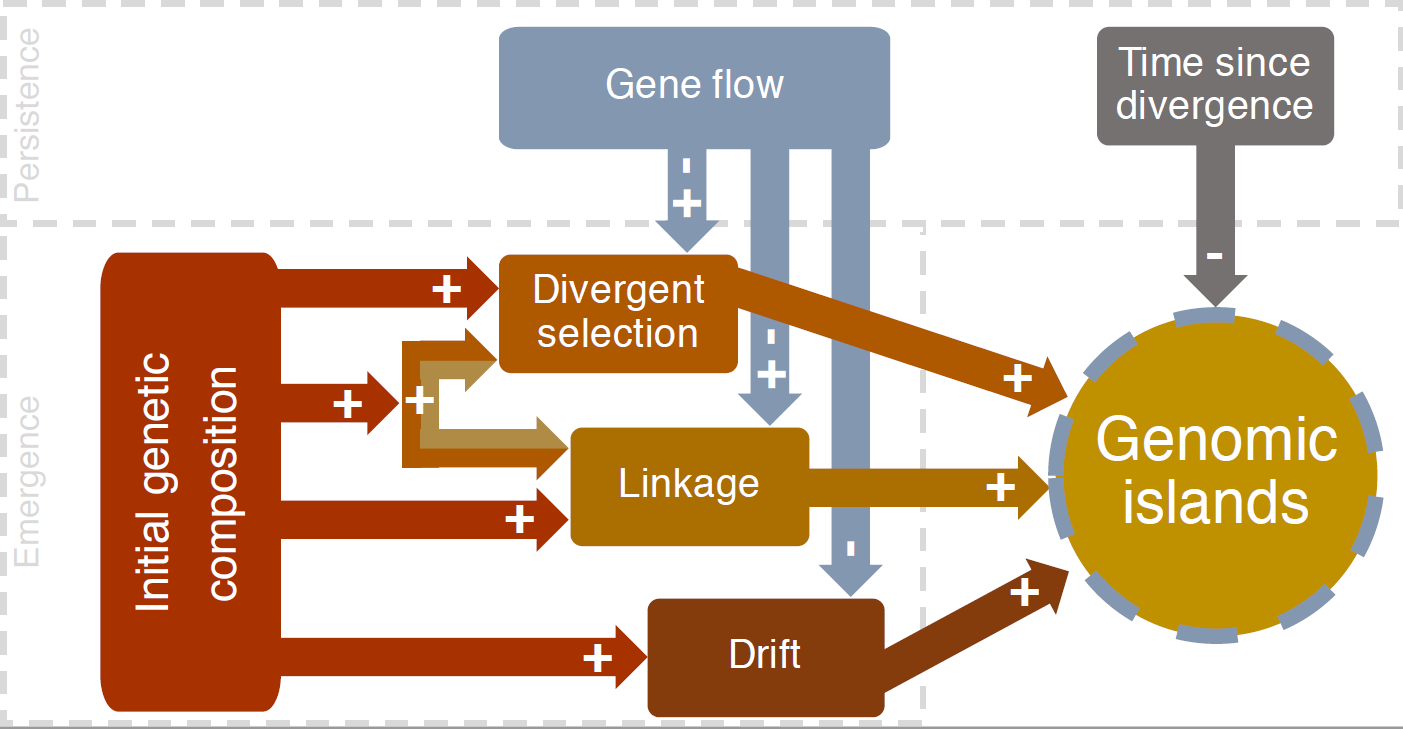
One striking pattern of genomic heterogeneity that can be explored with the ‘glads’ R package is that of genomic islands of divergence – highly differentiated peaks rising up out of a background of less differentiated regions across the genomes of divergent populations. The so-called metaphor characterises the genomic background as a sea, with peaks of divergence represented by islands, just like the one that our bird hatched in the Pacific.
Empirically, it was known that genomic islands of divergence could emerge under a range of different circumstances. One popular idea was they were the result of divergent selection acting on a portion of the genome, with surrounded areas homogenized by gene flow, hence the pattern was interpreted as evidence of divergence or speciation with ongoing gene flow. Yet, genomic islands were also observed in the absence of gene flow. On a songbird of the pacific, such as the one that hatched in our hypothetical case, divergence within and outside genomic islands has been shown to be important during the speciation process.
Through simulations, we investigated the range of conditions that are likely to result in the emergence of genomic islands of divergence. We demonstrated that high linkage disequilibrium, strong genetic drift, and strong divergent selection, could indeed result in the emergence of genomic islands. Yet, genomic island formation was not guaranteed because it also depended on genetic diversity of the starting population. The simulations also showed that genomic islands are more likely to be observed in recently diverged populations.
The role of gene flow in producing genomic islands had previously been heavily emphasised. We showed how different levels of gene flow could have a contrasting effect over time, slowing the rise of the surrounding sea by homogenising those genomic regions, but also acting to erase the pattern of islands under high levels of gene flow.
Summary
Our main conclusions stemming from our simulations of genomic islands of divergence are that interpreting a certain evolutionary process from a particular genomic pattern should be done cautiously. Yet, it may be possible to narrow the range of potential causes of a given genomic pattern through computational simulations. This would require additional eco-evolutionary information beyond genomic sequencing of a particular pair of divergent forms.
To find out more information about the R package ‘glads’ and the various processes influencing genomic islands of divergence, read our Methods in Ecology and Evolution manuscript “The many population genetic and demographic routes to islands of genomic divergence”.
Discover more about the articles that were shortlisted for this year’s Robert May Prize here.
